







































Вторинні посередники: цАМФ, цГМФ, інозитолтрифосфат та іони кальцію
Гормони та інші біологічно активні речовини(первинні месенджери) забезпечують передачу інформації між клітинами, але так як ці молекули знаходяться в внутрішньоклітинному середовищі, то для реалізації внутрішньоклітинних ефектів потрібні вторинні месенджери, які ми розлягнемо тут.

Введення
Клітини взаємодіють між собою за допомогою гормонів або інших біологічно активних речовин, які знаходяться у зовнішньоклітинному середовищі.На мембрані кожної клітини є якийсь рецептор, який зв’язується з певною молекулою, яку називають лігандом, цієї молекулою може бути будь-що, в тому числі гормони.Рецептор представляє собою трансмембранний білок, який пронизує мембрану навстіж, тобто, виступає як в міжклітинне середовище, так і в цитоплазму клітини.Таким чином, зовнішня частина рецептора може реагувати на зміни в зовнішньому середовищі, при цьому передаючи “сигнал” на внутрішню частину рецептора, яка знаходиться в цитоплазмі, внаслідок цього будуть активовуватись певні каскади реакцій - активація ферментів, факторів транскрипції, активація іонних каналів.
Так от, рецептори, які реагують на гормони, як правило являються зв’язаними з так званими G-білками(GTPase-protein), які складаються з трьох субодиниць:
- α субодиниця в неактивному стані з’єднана з молекулою ГДФ(гуанозиндифосфат)
- β субодиниця
- γ субодиниця
Є декілька видів G-білків:
- Gs-білок
- Gi-білок
- Gq-білок
Всі вони складаються з трьох субодиниць, але для кожного різновиду цього білку характерна різна активність α-субодиниці.Це можна уявити так: α-субодиниця - це шестерня, для Gs-білку характерна шестерня розміром 10 см, а для Gi-білку характерна шестерня 5 см, відповідно, вони мають різні властивості та запускають різні механізми.
Так от, коли якась сигнальна молекула зв’язується з рецептором, який зв’язаний з G-білком, то відбуваються конформаційні зміни в самому рецепторі. Які потім передаються на G-білок, в результаті чого α-субодиниця від’єднує ГДФ, а замість цього приєднує ГТФ, таким чином активуючись, і далі в залежності від типу активованого G-білку розвиваються різні реакції
Аденілатциклазний механізм
Якщо активовується α-субодиниця Gs-білку, то відбувається дисоціація цієї субодиниці від інших βγ субодиниць.α-субодиниця прямує в товщу клітинної мембрани, де знаходиться особливий фермент - аденілатциклаза.Коли активована субодиниця з’єднується з цим ферментом, то останній набуває каталітичної активності, починаючи перетворювати АТФ в циклічний аденозинмонофосфат(цАМФ).
В цитоплазмі клітини плаває фермент протеїнкіназа А, яка складається з чотирьох субодиниць:
- 2 регуляторні
- 2 каталітичні
Коли регуляторні субодиниці з’єднані з каталітичними, то останні не мають ніякої каталітичної активності.Циклічний АМФ(цАМФ) з’єднується з регулятоними субодиницями, стимулюючи їх дисоціацію від каталітичних, які тепер стають активними.
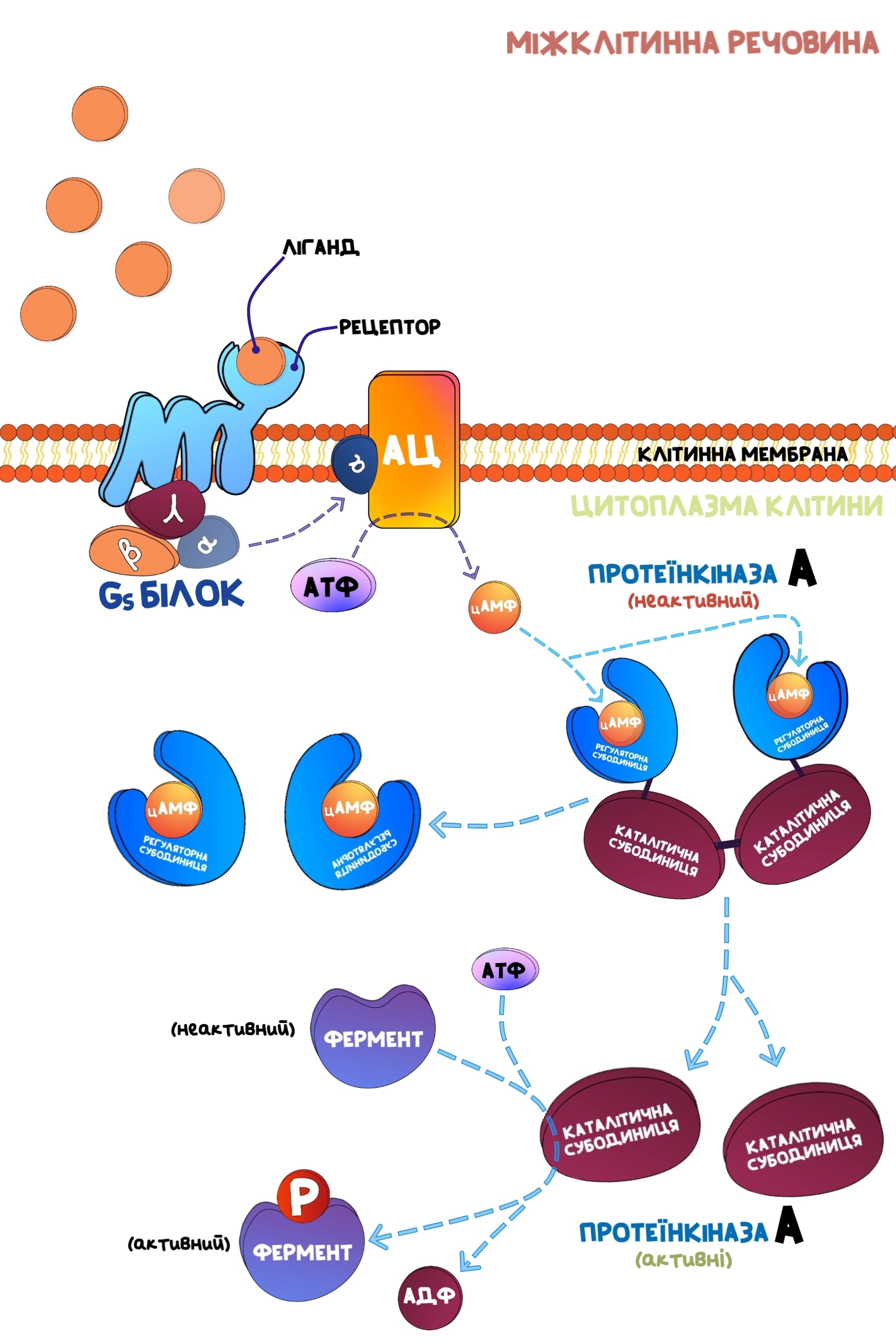
При зв'язувані ліганда(біологічно активної речовини, яка ідеально підходить для рецептора) з рецептором, який зв'язаний з G-білком, відбувається дисоціація α-субодиниці від інших, яка дифундує в товщу клітинної мембрани та активує аденілатциклазу.Останній активований фермент перетворює АТФ в цАМФ, який забезпечує процес відщеплення регуляторних субодиниць від каталітичних протеїнкінази А.
Каталітичні частини протеїнкінази А забезпечують протікання реакції фосфорилювання певних субстратів, якими в основному являються білки, ферменти та іонні канали.При цьому, донором фосфатної групи буде молекула АТФ, яка в результаті реакції перетворюється в АДФ.Фосфорилювання ферментів може як стимулювати його активність, так і пригнічувати, що саме буде відбуватись - залежить від ферменту.Тобто, протеїнкіназа А модулює активність певних білків(серед яких є ферменти).
Більше того, протеїнкіназа фосфорилює та таким чином активує фактор транскрипції CREBP(Cyclic-AMP Responsible Element Binding Protein), який після цього прямує в ядро та стимулює процес транскрипції певних генів.
Щоб не було сухої теорії, то наведу приклад: коли глюкагон зв’язується з відповідним рецептором на поверхні гепатоцита, то активується вище описаний каскад реакцій, в результаті чого протеїнкіназа А фосфорилює ферменту глікоген фосфорилазу, яка таким чином активується та починає розкладати глікоген на глюкозу, яка потім вийде в кров, таким чином проявляється гіперглікемічний ефект глюкагону.
Натомість, є ще Gi-білок, який при своїй активації також вивільняє α-субодиницю, яка в даному випадку буде не стимулювати, а пригнічувати аденілатциклазу, тобто, Gi- та Gs-білки являються антагоністами один відносно одного.
Інозитолтрифосфатний механізм
Якщо рецептор був зв’язаний з Gq-білком, то при активації α-субодиниці, вона знову ж таки дисоціює, прямує в товщу клітинної мембрани, але тепер активує тут фермент фосфоліпазу С.Активований фермент починає розщеплювати фосфатидилінозитол-4,5-дифосфат(структурний компонент клітинної мембрани) на диацилгліцерол та інозитол-1,4,5-трифосфат(ІФ3), останній в свою чергу прямує до гладкого ендоплазматичного ретикулуму(депо внутішньоклітинного кальцію), зв’язується з відповідним рецептором та стимулює викид кальцію з цієї органели в цитоплазму клітини.
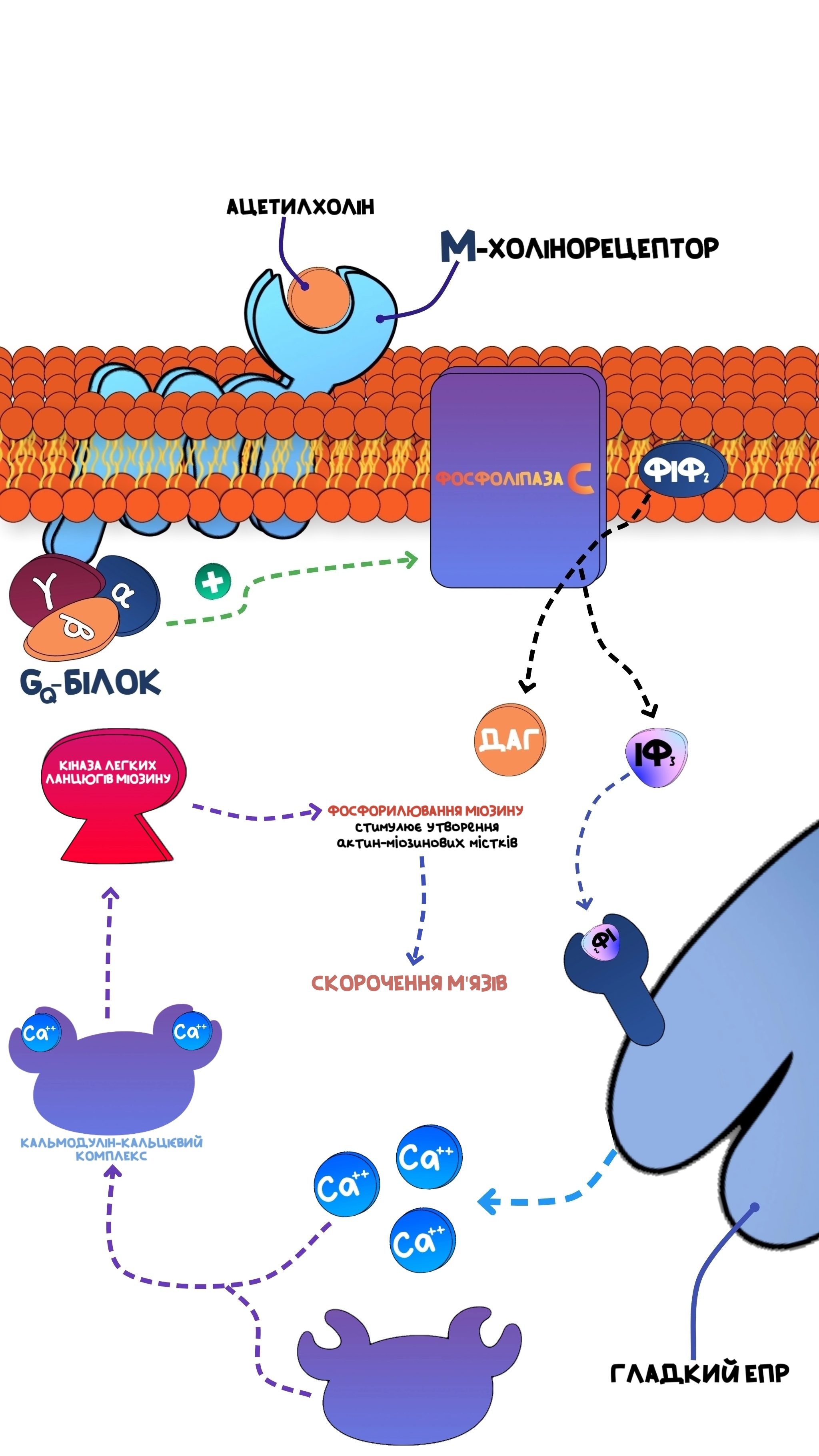
Знову ж таки, при активації рецептору(наприклад, М-холінорецептору) відбуваються конформаційні зміни всередині молекули Gq-білку, в результаті чого α-субодиниця дифундує в товщу клітинної мембрани, стимулює при цьому фосфоліпазу С, яка бере фосфатидилінозитол-4,5-дифосфат(компонент клітинної мембрани) та розщеплює його на ДАГ та ІФ3, останній стимулює виділення кальцію з внутрішньоклітинних депо(релевантно для гладких міоцитів).
ДАГ та ІФ3 разом забезпечують активацію протеїнкінази С, яка за аналогією до протеїнкінази А, здатна фосфорилювати, таким чином модулювати активність певних білків(протеїнкіназа А та протеїнкіназа С регулюють різні білки, це не потрібно плутати).За рахунок підвищення рівня внутрішньоклітинного кальцію відбувається скорочення гладких м’язів ШКТ, сечовивідних шляхів і т.д.
Гуанілатциклазний механізм
Ці реакції важливі для регуляції тонусу судин.
Внаслідок збільшення рівня кальцію(завдяки інозитолтрифосфатному механізму) в клітині ендотеліоцита відбувається активація кальмодуліну(кальцій-сенсорний білок), який перетворюється в фермент та набуває каталітичної активності.Після цього, за участю кальмодуліну відбувається активацію ендотеліальної нітроген оксид синтази(eNOS), яка використовує аргінін, розщеплює його на цитрулін та нітроген оксид(NO), останній являється газом, який вільно дифундує з клітини ендотеліоцита, потрапляє в гладкі міоцити стінки судини.В цитоплазмі будь-яких гладких міоцитів є розчинна форма гуанілатциклази, яка активується нітроген оксидом та починає перетворювати ГТФ(гуанозинтрифосфат) в цГМФ(циклічний гуанозинмонофосфат), а останній, в свою чергу, стимулює фермент протеїнкіназу Г, яка фосфорилює та таким чином модулює активність ферментів, які замішані в скорочення та розслаблення(фосфатаза легких ланцюгів міозину, кіназа легких ланцюгів міозину) гладкої м’язової клітини - відбувається розслаблення, і як наслідок знижується артеріальний тиск.
Хоча, розслаблення гладких м’язів може відбуватись і в результаті активації аденілатциклазного механізму(наприклад, в судинах, які кровопостачають скелетні м’язи).Загалом, циклічні мононуклеотиди сприяють розслабленню гладких м’язів.
Фосфодіестерази
Фосфодіестерази - ферменти, які перетворюють циклічні мононуклеотиди(цАМФ та цГМФ) в звичайні мононуклеотиди.Виділяють десь 15 таких ізоферментів, для певної тканини характерний такий-то ізофермент, але нам потрібно зрозуміти суть: якщо циклічних нуклеотидів в клітині накопичується дуже багато, то відбувається алостерична стимуляція цього ферменту, і він починає руйнувати циклічні нуклеотиди, щоб ті не накопичувались, таким чином регулюється нормальна функціональність клітини.Зокрема, в деяких випадках, цГМФ стимулює спеціальний ізофермент фосфодіестерази, і при активації цього ферменту, він буде руйнувати лише цАМФ, але не цГМФ, а може бути й навпаки.
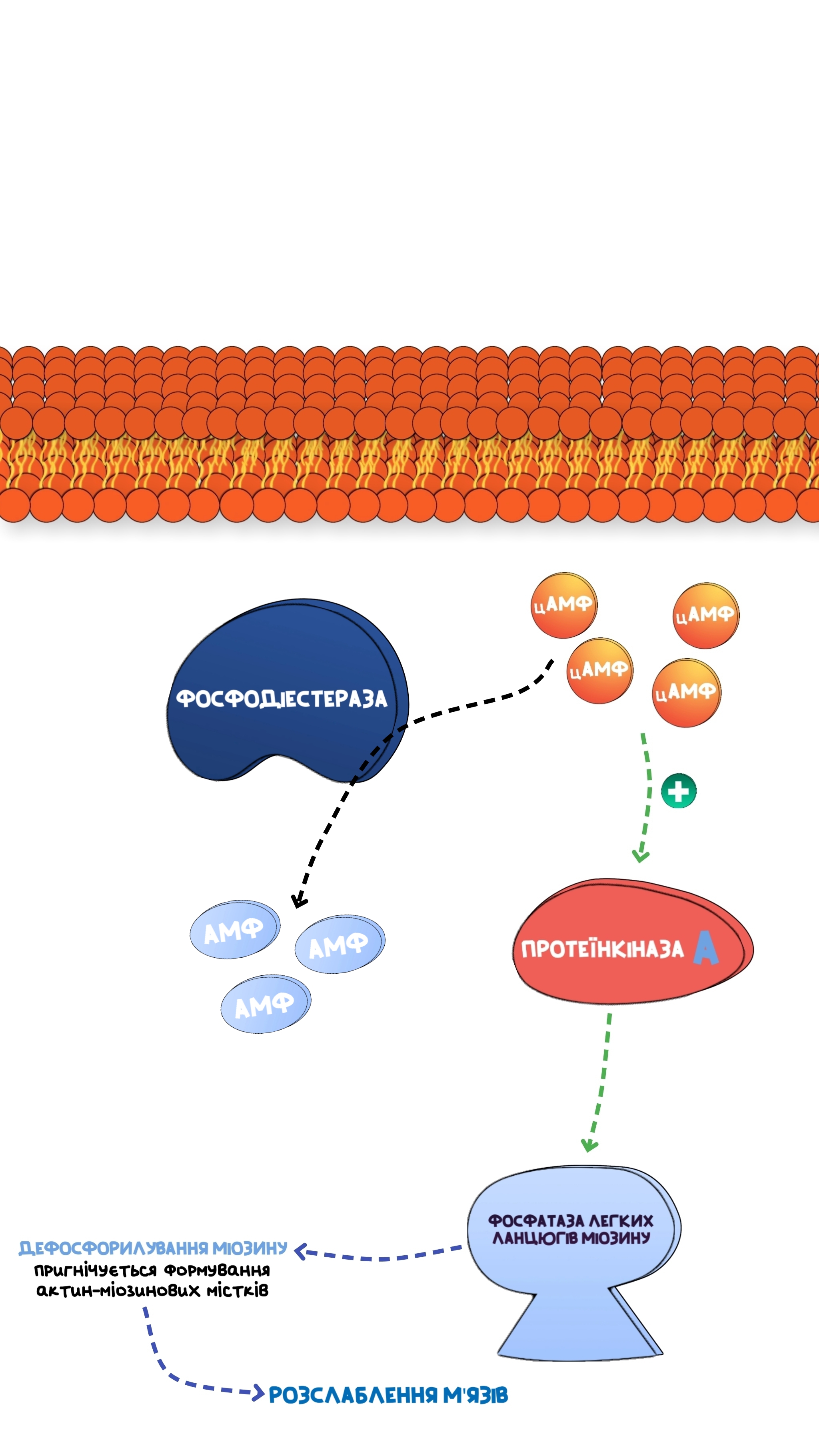
При накопичені циклічних мононуклеотидів в клітині, активується фосфодіестераза, яка каталізує перетворення циклічних в звичайні мононуклеотиди.
З приводу клінічного значення, то є такі фармакологічні препарати, які блокують фосфодіестеразу, відповідно, моноциклічні нуклеотиди будуть накопичуватись в клітині, що призведе до надмірної активності протеїнкіназ, які в свою чергу активують фосфатазу легких ланцюгів міозину, а вона дефосфорилює міозин, таким чином запобігає утворенню містків між цим білком та актином, відповідно, скорочення не відбувається, може використовуватись як спазмолітик(ношпа).



